

The Races of Humanity by Richard McCulloch
Divergent evolution is the process by which new life forms are created by the division and separation of life into different branches. Human evolution has seen its share of divergent branching. The generic name commonly used to refer to the genetically different populations, branches or divisions of humanity -- that share both a common biological ancestry and an ensemble of unique, genetically transmitted traits and characteristics which distinguish them from other populations -- is "race." But in the human species, as in any species enjoying a great degree of variety, the constant branching and dividing that characterizes the process of divergent evolution has created many different levels of branches and divisions, each of which possesses genetic traits which distinguish it from other branches or divisions at the same level. For purposes of taxonomic accuracy each of these levels should have its own specific name and definition. The first or highest level is called the species, and it is simply and objectively defined as including all those populations which are capable of interbreeding with each other and producing fully fertile offspring, and which do in fact interbreed under conditions of close and extensive contact. The term race is commonly used to refer to a branch or division of the species possessing genetically transmitted physical traits which distinguish it from other branches or divisions of the same level. Adding to this definition, it will here also be defined as including only those persons who are capable of reproduction with each other without the loss or significant diminishment or alteration of the racially-distinctive genetic traits of either parent stock. The genetically transmitted traits which distinguish a race from other divisions at the same level (i.e., other races) should not be diminished or lost by reproduction within the race. If racially-distinctive traits are lost or diminished by within-group reproduction then the population group is at a level of division too broad and inclusive to be accurately defined as a race. If it is too narrow to be defined as a species, as it does not include all those populations capable of interbreeding, then it is at a level between race and species, which will here be referred to as a subspecies.
The closest living relative of humanity, the still-existing species most closely related to Homo sapiens, is the Chimpanzee, whose ancestral line separated and branched from the line leading to humans about 5.5 million years ago. Even after 5.5 million years of divergent evolution humans and chimpanzees still have over 98% of their genes in common, with only a 1.23% (Time, October 9, 2006) to 1.6% difference in their genome. The genus Homo originated with Homo habilis in the region of the Great Rift Valley in Kenya and Ethiopia in east Africa about two million years ago, where it continued to evolve, first as Homo ergasterand Homo erectus, then as Homo antecessor (750,000 years ago) and Homo heidelbergensis (600,000-250,000 years ago; believed to be the direct ancestor of Homo neanderthalensis in western Eurasia), and then as Homo sapiens idaltu, the earliest modern humans, with finds in Ethiopia dated to 195,000 and 160,000 years ago (Scientific American, Vol. 16, No. 2, 2006, p. 78).
It is generally agreed that there were at least three major migrations or expansions of the genus Homo out of east Africa into Eurasia, either crossing the Sinai peninsula from Egypt into the Levant (the coast of what is now Israel, Lebanon and Syria), or crossing the southern entrance of the Red Sea (the Bab el Mandeb) from Djibouti in Africa to Yemen in Asia, from where they spread throughout most of Eurasia and developed into a variety of regional "archaic" human populations. The first of these major expansions out of east Africa into Eurasia was about 1.8 million years ago, the second about 600,000 years ago (associated with the spread of the Acheulean culture), and the last shortly after 100,000 years ago. Beginning in 1987, based on genetic studies showing that the mitochondrial DNA (mtDNA) and the Y-chromosome of all living humans is derived from the last of these major expansions, the common view expressed in the popular press (called "Out-of-Africa") has been that the modern humans of the final migration completely replaced the regional archaic human populations from the first two major expansions. But beginning in 2002 studies of other genes by Alan Templeton and others have increasingly supported the view (called "Out-of-Africa-Again-and-Again") that although all our surviving mtDNA and Y-chromosome lineages as well as the majority of our other genes derive from the most recent expansion, a significant minority of our other genes have much older "coalescence" dates and must therefore derive from the regional archaic human populations of the first two major expansions. These studies indicate that some genes from the regional populations of the first expansion were assimilated and perpetuated by the populations of the second expansion, and that some of the genes of both of the first two (archaic) expansions were assimilated by the modern humans of the final expansion.
The first dispersal of modern humans probably began soon after the emergence of Homo sapiens idaltu in east Africa about 195,000 years ago, with some populations heading west into the tropical forest of the Congo basin where they evolved into the Congoid subspecies (possibly with the assimilation of some local archaic elements), others remaining in east Africa where they evolved into the Capoid or Khoisanid (San-Bushmen) subspecies, and others moving north to the shores of the Red Sea, where they became the progenitors of the population that eventually migrated out of Africa and populated the rest of the world, possibly assimilating some of the regional archaic human populations they encountered in varying degrees, and evolving into the Australoid, Mongoloid and Caucasoid subspecies. By 130,000 years ago there were perhaps 10,000 modern humans living in different populations in different regions of Africa. About 120,000 years ago one of these modern human populations that had expanded up the Nile valley crossed the Sinai peninsula out of Africa into the Levant but got no further, and by 90,000 years ago its members had either returned to Africa or died out.
The following account of the final major expansion out of east Africa into Eurasia, that of the modern humans shortly after 100,000 years ago, is based largely on the work of Stephen Oppenheimer as detailed in his book Out of Eden: The Peopling of the World (2004) which was also the basis for a Discovery Channel documentary titled The Real Eve.
The migrating modern human population, probably numbering only a few hundred people at the beginning, crossed from Africa to Asia at the southern entrance of the Red Sea. From there they followed a beachcombing trek that took them along the coastline of the Arabian Sea. The descendants of this population gradually expanded and dispersed, with the initial expansion being along the southern coast of Asia. The where and when of these early human migrations was largely determined by geography, especially changes in climate and sea level. The first main split or division in the expansion occurred on the Iranian coast of the Persian Gulf, with some groups continuing to move east while others remained in southern Iran between the Zagros Mountains and the sea. The second main branching or division probably occurred in southeast Asia, with one group continuing to move eastward, reaching China by 68,000 years ago, and another group remaining in the Burma-Thailand region where it evolved into a proto-Australoid population and then expanded south through Malaysia and Indonesia, reaching New Guinea by 77,000 years ago and Australia by 65,000 years ago.
The eruption, or explosion, of the Toba super-volcano in northern Sumatra circa 74,000 years ago, the largest such explosion in the last two million years, perhaps 100 times larger than the Krakatoa event off southern Sumatra in 1883, covered the entire Indian sub-continent in several meters of ash, probably destroying almost all life, including the early human population in the area. The populations to the east and south of the eruption were spared its catastrophic effects, but the population in southern Iran, and to a lesser extent the population in east Africa, probably suffered severe climate effects. The population in west Africa, protected by mountains to the east, was not as seriously effected. Within a few thousand years India was repopulated from the east by proto-Australoids.
By 50,000 years ago the population that had remained in southern Iran had evolved into proto-Caucasoids and began to expand -- to the east into Pakistan and northern India; to the northwest up the Tigris-Euphrates valley to the Levant by 45,000 years ago; and to the northeast through Central Asia to Russia and the steppes of western Siberia, also by 45,000 years ago. From the Levant they expanded north into Anatolia, from there entering Europe through the Balkans and spreading the Aurignacian culture across southern Europe by 43,000 years ago. From Russia they moved westward into Europe, spreading theGravettian culture, about 33,500 years ago. Shortly after this another Caucasoid group expanded from the Levant across North Africa. In this same time frame the population in Indochina and southern China had evolved into proto-Mongoloids and expanded northwards into the steppes of eastern Siberia, branching into southern and northern Mongoloid groups. Some northern Mongoloids migrated northeast to Berengia, a vast land between Siberia and Alaska that is now underwater, from where they subsequently moved south into the Americas.
By 30,000 years ago the divergent evolutionary branching or dividing of the human species had produced five main lines or subspecies which are still extant -- the Congoid of West Africa; the Capoid of East and South Africa (later replaced in East Africa by the Congoid); the Australoid of India, Burma, Malaya, Indonesia, New Guinea and Australia; the Mongoloid of East Asia (later expanding to the southwest into Burma, Malaya and Indonesia, largely replacing the indigenous Australoids) and the Caucasoid of Europe, North Africa and West Asia (partly replacing the Mongoloids in the Americas after A.D. 1492 and the Australoids in Australia after A.D. 1788). These subspecies branched or divided in turn into separate races, and these races branched in their turn into subraces, as part of the continuing process of divergent evolution.
Beginning about 20,000 years ago, when the global human population was perhaps a million, the Last Glacial Maximum (LGM) pushed the population of northern Europe south to refuge areas in southern France, northern Spain, the Balkans and Ukraine, while the now fully-developed northern Mongoloid population in Siberia was also forced south to eastern and southern China. Both populations were greatly reduced in number during this period. (The expansion of the southern Mongoloids into Malaya and Indonesia, partly replacing and partly assimilating the native Australoids, probably occurred during this period.) When the Last Glacial Maximum began to recede about 15,000 years ago (13,000 B.C.) the survivors of these populations expanded northward again from their refuge areas, with Scandinavia being occupied by humans for the first time about 10,000 years ago, by which time the global human population had risen to about 10 million. Agriculture and the Neolithic period also began about 10,000 years ago in both the Middle East and China. The genetic ancestry of the native European population as a whole is about 80% from the original Upper Paleolithic inhabitants who survived the 5,000 years of the Last Glacial Maximum in southern refuge areas and then re-expanded and repopulated the central and northern regions of the continent, and 20% from the Neolithic farmers who expanded from Anatolia into Europe starting about 8,000 years ago, with the latter element concentrated primarily in the Mediterranean lands of southern Europe, indicating that the initial spread of agriculture into central and northern Europe was a process of cultural diffusion rather than a movement of people.
The different races are often popularly defined and named (often inaccurately) by skin color, but as this system is based on only one genetic phenotypic difference, when hundreds are involved, it tends to distort the reality of race and racial differences. In the system of racial classification outlined below the names assigned to the various subspecies and races are, with a few exceptions, based on geographical regions where they are the native type.
- Outline of Human Racial Classification:
- SUB-SAHARAN AFRICA GROUP
- I. Capoid or Khoisanid Subspecies of southern Africa
- A. Khoid (Hottentot) race
- B. Sanid (Bushmen) race
- II. Congoid Subspecies of sub-Saharan Africa
- A. Central Congoid race (Geographic center and origin in the Congo river basin)
- 1. Palaecongoid subrace (the Congo river basin: Ivory Coast, Ghana, Nigeria, Cameroon, Congo, Angola)
- 2. Sudanid subrace (western Africa: Niger, Mali, Senegal, Guinea)
- 3. Nilotid subrace (southern Sudan; the ancient Nubians were of this subrace)
- 4. Kafrid or Bantid subrace (east and south Africa: Kenya, Tanzania, Mozambique, Natal)
- B. Bambutid race (African Pygmies)
- C. Aethiopid race (Ethiopia, Somalia; hybridized with Caucasoids)
- "OUT-OF-AFRICA" GROUP
- I. Australoid Subspecies
- A. Veddoid race (remnant Australoid population in central and southern India)
- B. Negritos (remnants in Malaysia and the Philippines)
- C. Melanesian race (New Guinea, Papua, Solomon Islands)
- D. Australian-Tasmanian race (Australian Aborigines)
- II. Mongoloid Subspecies
- A. Northern Mongoloid racial group
- 1. Northeast Asian race (various subraces in northern China, Manchuria, Korea and Japan)
- 2. Ainuid race (remnants of aboriginal population in northern Japan)
- 3. Tungid race (Mongolia and Siberia, Eskimos)
- 4. Amerindian race (American Indians; various subraces)
- B. Southern Mongoloid racial group
- 1. Southeast Asian race (various subraces in southern China, Indochina, Thailand, Myanmar [Burma], Malaysia, Indonesia and the Philippines, the last four partly hybridized with Australoids)
- 2. Micronesian-Polynesian race (predominantly Southern Mongoloid partly hybridized with Australoids)
- III. Caucasoid or Europid Subspecies
- A. Dravidic race (India, Bangladesh and Sri Lanka [Ceylon]; ancient stabilized Indic-Veddoid [Australoid] blend)
- B. Turanid race (partially hybridized with Mongoloids; predominant element in Kazakhstan, Turkmenistan and Uzbekistan; common in Hungary and Turkey)
- C. Indic or Nordindid race (Pakistan and northern India)
- D. Irano-Afghan race (predominant in Iran and Afghanistan, primary element in Iraq, common [25%] in Turkey)
- E. Armenid race (predominant element in Armenia and Azerbaijan, common in Syria, Lebanon and northern Iraq, primary element among the Ashkenazic Jews)
- F. Mediterranid racial group
- 1. Orientalid or Arabid subrace (predominant in Arabia, major element from Egypt to Syria, primary in northern Sudan, important in Iraq, predominant element among the Oriental Jews)
- 2. South Mediterranean or Saharid subrace (predominant in Algeria and Libya, important in Morocco, Tunisia and Egypt, primary element among the Sephardic Jews, common element [circa 20-25%] in Spain, Sicily and southern Italy, minor element [circa 5%] in Greece)
- 3. East Mediterranean or Pontid subrace (Black Sea coast of Ukraine, Romania and Bulgaria; Aegean coasts of Greece and Turkey)
- 4. Dinaricized Mediterraneans (Residual mixed types resulting from the blending of Mediterranids with Dinarics, Alpines or Armenids; not a unified type, has much regional variation; predominant element [over 60%] in Sicily and southern Italy, principal element in Turkey [35%], important element in western Syria, Lebanon and central Italy, common in northern Italy. The ancient Cappadocian Mediterranean subrace of Anatolia was dinaricized perhaps as early as the Neolitihic and is a major contributor to this type in modern Turkey.)
- 5. West Mediterranean or Iberid subrace (Spain, Portugal, Corsica, Sardinia, and coastal areas of Morocco and Tunisia; the Atlanto-Mediterranean peoples who expanded over much of the Atlantic coastal regions of Europe during the Mesolithic period were a branch of this subrace)
- G. Ladogan race (named after Lake Ladoga; indigenous to Russia; includes Lappish subrace of arctic Europe)
- H. Dinaric race (predominant in western Balkans [Dinaric Mountains] and northern Italy, important in the Czech Republic, eastern and southern Switzerland, western Austria and eastern Ukraine. Its distribution in Europe, and that of its derived Dinaricized Mediterranean type, may be associated with the expansion of the Neolithic Anatolian farmers beginning circa 6,500 B.C.)
- I. Alpine race (predominant element in Luxembourg, primary in Bavaria and the Czech Republic [Bohemia], important in France, Hungary, eastern and southern Switzerland)
- J. Nordish or Northern European racial group (various subraces in the British Isles, Scandinavia, the Netherlands and Belgium; predominant element in Germany, Switzerland, Poland, Finland and the Baltic States; majority in Austria and Russia; important in France, the Czech Republic, Slovakia and Hungary; outlined in detail in The Nordish Race)
- Dominant or predominant = over 60% majority
- Majority or major = 50-60% majority
- Principal or primary = 25-49% plurality; less than a majority, but most numerous racial type
- Important = 25-49% minority; not most numerous racial type
- Common = 6-25% minority
- Minor = 5% or less minority
What are the percentages of genetic differences between the human races? Perhaps the best study to date (2010) on this subject is still that of Masatoshi Nei and Arun K. Roychoudhury from Evolutionary Relationships of Human Populations on a Global Scale (1993). The following table (Fig. 1 below) of estimates of genetic differences between human populations is from their study.
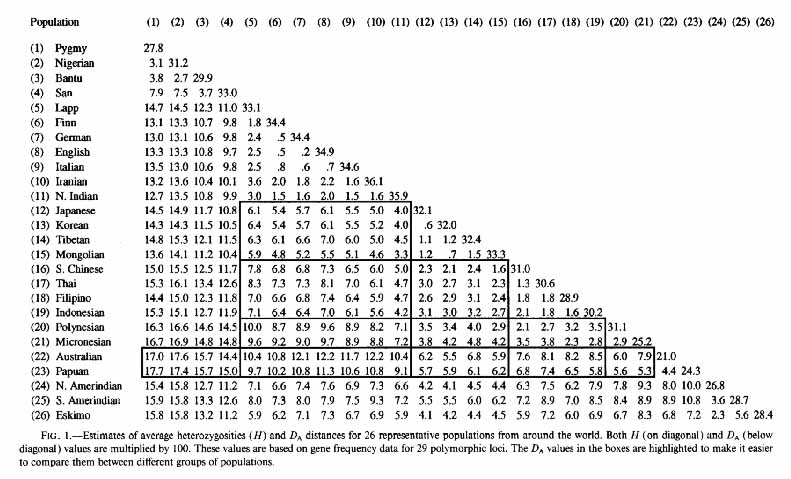
The following table of percentages of genetic differences between human populations presents the estimates for 19 populations from the above table in an easier to read and understand format. The human-chimpanzee genetic difference, giving the greatest degree of difference from the commonly accepted range using the same methodology, is added for context and comparison.
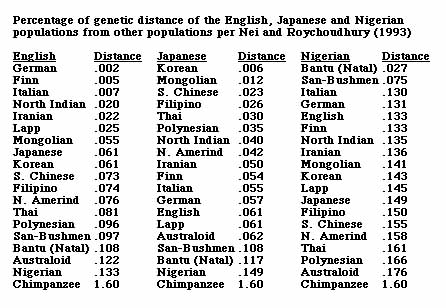
If one were to spatially visualize the first column of the above scale, with a German standing at a distance of 20 feet from an Englishman, a Finn would stand at a distance of 50 feet, an Italian at 70 feet, a northern Indian at 200 feet, a Japanese at 610 feet, a North American Amerindian at 760 feet, a Nigerian at 1,330 feet, and a Chimpanzee at 16,000 feet. The greatest percentage of genetic difference is .176% between Nigerians and Australian Aborigines. This is 11% of the genetic difference of 1.6% between humans and chimpanzees, different biological Families whose ancestral lines are believed to have separated 5-7 million years ago. The .133% genetic difference between the English and Nigerian populations is 8.3% as large as the genetic difference between humans and chimpanzees. The .061% genetic difference between the English and Japanese or Korean populations is 3.8% as large as the genetic difference between humans and chimpanzees. Seen in this context, these are very significant genetic differences. It is also worth noting that for both the English and the Japanese, representing Europeans and Northeast Asians, the greatest percentage of genetic difference is with the Nigerians, and that the degree of this difference, .133% for the English and .149% for the Japanese, is very similar. By comparison, the English and Japanese degree of difference from the Australian Aborigine population, .122% for the English and .062% for the Japanese, is very different, with the English-Australoid difference twice as great as the Japanese-Australoid difference.
Return to Racial Compact main page
No comments:
Post a Comment